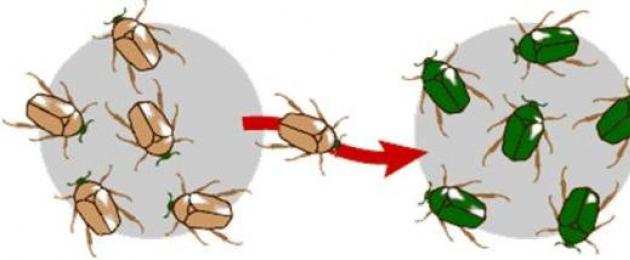
Участок ДНК, на котором размещается определенный ген, называется локусом. В нем могут содержаться альтернативные варианты генетической информации - аллели. В любой популяции есть большое количество данных структур. При этом доля конкретного аллеля в общем геноме популяции носит название частоты гена.
Чтобы определенная мутация привела к эволюционным изменениям видов, ее частота должна быть достаточно высокой, а мутантный аллель должен фиксироваться во всех индивидуумах каждого поколения. При незначительном ее количестве мутационные изменения не способны повлиять на эволюционную историю организмов.
Чтобы частота аллеля росла, должны действовать определенные факторы - дрейф генов, миграция и
Дрейф генов - это случайный рост аллеля при воздействии нескольких событий, которые сочетаются и имеют стохастический характер. Данный процесс связывается с тем, что не все лица в популяции принимают участие в размножении. Он наиболее характерен для признаков или заболеваний, которые встречаются редко, но вследствие отсутствия отбора способны храниться в роду или даже целой популяции небольшого размера в течение длительного времени. Такая закономерность часто прослеживается в малой которой не превышает 1000 особей, поскольку в данном случае чрезвычайно мала миграция.
Для того чтобы лучше понять дрейф генов, следует знать следующие закономерности. В случаях, когда частота аллеля составляет 0, в последующих поколениях она не меняется. Если же она достигает 1, то говорят, что ген в популяции фиксируется. Случайный дрейф генов и является следствием процесса фиксации при одновременной потере одного аллеля. Чаще всего данная закономерность прослеживается тогда, когда мутации и миграции не вызывают постоянного изменения составляющих локусов.

Поскольку частота генов имеет ненаправленный характер, она уменьшает разнообразие видов, а также увеличивает различия между локальными популяциями. Стоит отметить, что этому противодействует миграция, при которой разные группировки организмов обмениваются своими аллелями. Надо также сказать, что дрейф генов практически не влияет на частоту отдельных генов в больших популяциях, но в он может стать решающим При этом количество аллелей резко меняется. Некоторые гены могут безвозвратно теряться, что значительно обедняет генетическое разнообразие.

В качестве примера можно привести массовые эпидемии, после которых восстановление популяции проводилось практически за счет нескольких ее представителей. При этом все потомки имели идентичный предкам геном. В дальнейшем расширение аллельного многообразия обеспечивалось завозом производителей или выездными вязками, которые способствуют росту различий на генном уровне.
Крайним проявлением дрейфа генов можно назвать появление совершенно новой популяции, которая образуется только от нескольких особей - так называемый эффект основателя.
Следует сказать, что закономерности перестройки генома изучает биотехнология. - это методика данной науки, которая позволяет переносить наследственную информацию. При этом перенос генов позволяет бороться с межвидовым барьером, а также придавать организмам необходимые свойства.
«дрейфуют» независимо. Поэтому результаты дрейфа оказываются разными в разных популяциях - в одних фиксируется один набор аллелей, в других - другой. Таким образом, дрейф генов ведет с одной стороны к уменьшению генетического разнообразия внутри популяций, а с другой стороны - к увеличению различий между популяциями, к их дивергенции по ряду признаков. Эта дивергенция в свою очередь может служить основой для видообразования.
В ходе эволюции популяций дрейф генов взаимодействует с другими факторами эволюции, прежде всего с естественным отбором . Соотношение вкладов этих двух факторов зависит как от интенсивности отбора, так и от численности популяций. При высокой интенсивности отбора и высокой численности популяций влияние случайных процессов на динамику частот генов в популяциях становится пренебрежимо малым. Наоборот, в малых популяциях при небольших различиях по приспособленности между генотипами дрейф генов приобретает решающее значение. В таких ситуациях менее адаптивный аллель может зафиксироваться в популяции, а более адаптивный может быть утрачен.
Как мы уже знаем, наиболее частым последствием дрейфа генов является обеднение генетического разнообразия внутри популяций за счет фиксации одних аллелей и утраты других. Мутационный процесс, напротив, приводит к обогащению генетического разнообразия внутри популяций. Аллель, утраченный в результате дрейфа, может возникать вновь и вновь за счет мутирования.
Поскольку дрейф генов - ненаправленный процесс, то одновременно с уменьшением разнообразия внутри популяций, он увеличивает различия между локальными популяциями. Этому противодействует миграция. Если в одной популяции зафиксирован аллельА , а в другой а , то миграция особей между этими популяциями приводит к тому, что внутри обеих популяций вновь возникает аллельное разнообразие.

Рис. 3. N - число особей в популяции. Видно, что при 25 особях после 40-го поколения один аллель исчезает, при 250 - соотношение аллелей меняется, а при 2500 - остается близким к исходному.
Эффект бутылочного горлышка сыграл, по-видимому, очень значительную роль в эволюции популяций человека. Предки современных людей в течение десятков тысяч лет расселялись по всему миру. На этом пути, множество популяций полностью вымирало. Даже те, которые уцелели, часто оказывались на грани вымирания. Их численность падала до критического уровня. Во время прохождения через «бутылочное горлышко» численности частоты аллелей менялись по-разному в разных популяциях. Определенные аллели утрачивались полностью в одних популяциях и фиксировались в других. После восстановления численности популяций их измененная генетическая структура воспроизводилась из поколения в поколение. Эти процессы, по-видимому, и обусловили, то мозаичное распределение некоторых аллелей, которое мы сегодня наблюдаем в локальных популяциях человека. Ниже представлено распределение аллеляВ по системе групп крови АВ0 у людей. Значительные отличия современных популяций друг от друга могут отражать последствия дрейфа генов, который происходил в доисторические времена в моменты прохождения предковых популяций через «бутылочное горлышко» численности.
Генетико-автоматические процессы, или дрейф генов, приводят к сглаживанию изменчивости внутри группы и появлению случайных, не связанных с отбором различий между изолятами. Именно это выявили наблюдения за особенностями фенотипов малочисленных групп населения в условиях, например, географической изоляции. Так, среди жителей Памира резус-отрицательные индивидуумы встречаются в 2—3 раза реже, чем в Европе. В большинстве кишлаков такие люди составляют 3—5% популяции. В некоторых изолированных селениях, однако, их насчитывается до 15%, т.е. примерно как в европейской популяции.
В крови человека имеются гаптоглобины , которые связывают свободный гемоглобин после разрушения эритроцитов, чем предотвращают его выведение из организма. Синтез гаптоглобина Нр1-1 контролируется геном Нр1. Частота этого гена у представителей двух соседних племен на Севере Южной Америки составляет 0,205 и 0,895, отличаясь более чем в 4 раза.
Примером действия дрейфа генов в человеческих популяциях служит эффект родоначальника. Он возникает, когда несколько семей порывают с родительской популяцией и создают новую на другой территории. Такая популяция обычно поддерживает высокий уровень брачной изоляции. Это способствует случайному закреплению в ее генофонде одних аллелей и утрате других. В результате частота очень редкогоаллеля может стать значительной.
Так, члены секты амишей в округе Ланкастер штата Пенсильвания, насчитывающей к середине девятнадцатого века примерно 8000 человек, почти все произошли от трех супружеских пар, иммигрировавших в Америку в 1770 г. В этом изоляте обнаружено 55 случаев особой формы карликовости с многопалостью, которая наследуется по аутосомно-рецессивному типу. Эта аномалия не зарегистирирована среди амишей штатов Огайо и Индиана. В мировой медицинской литературе описано едва ли 50 таких случаев. Очевидно, среди членов первых трех семей, основавших популяцию, находился носитель соответствующего рецессивного мутантного аллеля — «родоначальник» соответствующего фенотипа.
В XVIII в. из Германии в США иммигрировало 27 семей, основавших в штате Пенсильвания секту дункеров. За 200-летний период существования в условиях сильной брачной изоляции генофонд популяции дункеров изменился в сравнении с генофондом населения Рейнской области Германии, из которой они произошли. При этом степень различий во времени увеличивалась. У лиц в возрасте 55 лет и выше частоты аллелей системы групп крови MN ближе к цифрам, типичным для населения Рейнской области, чем у лиц в возрасте 28—55 лет. В возрастной группе 3—27 лет сдвиг достигает еще больших значений (табл. 1).
Рост среди дункеров лиц с группой крови М и снижение — с группой крови N нельзя объяснить действием отбора, так как направление изменений не совпадает с таковым в целом для населения штата Пенсильвания. В пользу дрейфа генов говорит также то, что в генофонде американских дункеров увеличилась концентрация аллелей, контролирующих развитие заведомо биологически нейтральных признаков, например оволосения средней фаланги пальцев, способности отставлять большой палец кисти (рис. 4).
Таблица 1. Прогрессивное изменение концентрации аллелей системы групп крови MN в популяции дункеров
На протяжении большей части истории человечества дрейф генов оказывал влияние на генофонды популяций людей. Так, многие особенности узкоместных типов в пределах арктической, байкальской, центрально-азиатской, уральской групп населения Сибири являются, по-видимому, результатом генетико-автоматических процессов в условиях изоляции малочисленных коллективов. Эти процессы, однако, не имели решающего значения в эволюции человека.
Рис. 4. Распространение нейтральных признаков в изолятедункеров штата Пенсильвания: а— рост волос на средней фаланге пальцев кисти, б— способность отставлять большой палец кисти
Последствия дрейфа генов, представляющие интерес для медицины, заключаются в неравномерном распределении по группам населения Земного шара некоторых наследственных заболеваний. Так, изоляцией и дрейфом генов объясняется, по-видимому, относительно высокая частота церебромакулярной дегенерации 1 в Квебеке и Ньюфаундленде, детского цестиноза во Франции , алкаптонурии в Чехии, одного из типов порфирии среди европеоидного населения в Южной Америке, адреногенитального синдрома у эскимосов. Эти же факторы могли быть причиной низкой частоты фенилкетонурии у финнов и евреев-ашкенази.
Изменение генетического состава популяции вследствие генетико-автоматических процессов приводит к гомозиготизации индивидуумов . При этом чаще фенотипические последствия оказываются неблагоприятными. Гомозиготизация - это перевод гетерозигот в гомозиготы при близкородствснном скрещивании. Ч. Дарвин описывает явление, которое вполне можно объяснить дрейфом генов. «Кролики, одичавшие на острове Порто-Санто, близ о. Мадейры», заслуживают более полного описания*. Вместе с тем следует помнить, что возможно образование и благоприятных комбинаций аллелей. В качестве примера рассмотрим родословные Тутанхамона (рис.5) и Клеопатры VII (рис. 6), в которых близкородственные браки были правилом на протяжении многих поколений.
Тутанхамон умер в возрасте 18 лет. Анализ его изображения в детском возрасте и подписи к этому изображению позволяют предположить, что он страдал генетическим заболеванием — целиакией , которая проявляется в изменении слизистой оболочки кишечника, исключающем всасывание клейковины.
________________________________________________________
1 церебромакулярная дегенерация, болезнь Тея - Сакса. Относится к группе наследственных липидных болезней мозга. На основании возраста начала болезни, клинических проявлений, картины глазного дна и данных биохимического исследования выделяют 5 форм амавротическойидиотии: врожденную, раннюю детскую, позднюю детскую, ювенильную и позднюю. Некоторые из этих форм отличаются и по характеру наследования.Характерный признак заболевания - диффузная дегенерация ганглиозных клеток во всех отделах нервной системы. Процесс распада ганглиозных клеток и превращения многих из них в зернистую массу - шафферовская дегенерация - является патогномоничным признаком амавротическойидиотии. Отмечаются также распад миелиновых волокон, особенно в зрительных и пирамидных путях, дегенеративные изменения глии.Врожденная форма - редкое заболевание. У ребенка уже при рождении отмечается микро- или гидроцефалия, параличи, судороги. Быстро наступает смерть. В мозговой ткани увеличено содержание ганглиозида Gm3.
Тутанхамон родился от брака Аменофиса III и Синтамоне, которая была дочерью Аменофиса III. Таким образом, мать фараона была его сводной сестрой. В могильном склепе Тутанхамона обнаружены мумии двух, по всей видимости мертворожденных, детей от брака с Анкесенамон, его племянницей.
Первая жена фараона была или его сестрой, или дочерью. Брат Тутанхамона Аменофис IV предположительно страдал болезнью Фрелиха и умер в 25—26 лет. Его дети от браков с Нефертити и Анкесенамон (его дочерью) были бесплодны. С другой стороны, известная своим умом и красотой Клеопатра VII была рождена в браке сына Птоломея Х и его родной сестры, которому предшествовали кровнородственные браки на протяжении по крайней мере шести поколений.
________________________________________________________________
*Это интересно
В 1418 или 1419 г. у ГонзалесаЗарко на корабле случайно оказалась беременная крольчиха, которая родила во время путешествия. Все детеныши были выпущены на остров. Кролики уменьшились почти на три дюйма в длину и почти вдвое в весе тела. По окраске кролик с Порто-Санто значительно отличается от обыкновенного. Они необычайно дики и проворны. По своим привычкам они более ночные животные. Производят от 4 до 6 детенышей в помете. Не удалось спарить с самками других пород". Примером воздействия дрейфа генов могут быть кошки о. Вознесения. Более 100 лет назад на острове появились крысы. Они расплодились в таком количестве, что английский комендант решил избавиться от них с помощью кошек. По его просьбе привезли кошек. Но они сбежали в отдаленные уголки острова и стали уничтожать не крыс, а домашнюю птицу и диких цесарок.
Другой комендант, чтобы избавиться от кошек, завез собак. Собаки не прижились — они ранили лапы об острые кромки шлака. Кошки со временем стали свирепыми и кровожадными. За столетие они отрастили себе почти собачьи клыки и стали сторожить дома островитян, ходить по пятам за хозяином и бросаться на посторонних.
Рис. 5. Родословная фараона XVIII династии Тутанхамона
Рис. 6. Родословная Клеопатры VII
Заключение и выводы:
Традиционно волны численности (жизни, популяционные) — присущие всем видам периодические и апериодические изменения численности особей в результате влияния абиотических и биотических факторов, воздействующих на популяцию, считаются "поставщиком" элементарного эволюционного материала.
Наилучшим доказательством значения дрейфа генов в микроэволюции
служит характер случайной локальной дифференциации в серии перманентноили периодически изолированных маленьких колоний. Дифференциация подобного типа многократно обнаруживалась в различных группах животных ирастений, популяции которых представляют собой систему колоний. Этадифференциация, если и не доказывает, то по крайней мере сильно склоняет кмнению о том, что дрейф генов играет важную роль в популяционных системах такого типа.
Использованная литература:
1. Гинтер Е.К Медицинская генетика: Учебник. - М.: Медицина, 2003. - 448 с.: ил
2. Грин Н., Стаут У., Тейлор Д «Биология» в 3 томах Москва «Мир» 2000г
3. Гуттман Б., Гриффитс Э., Сузуки Д., Кулис Т. Генетика. М.: ФАИР - ПРЕСС, 2004., 448 с
4. Жимулев И.Ф Генетика. Издательство Сибирского университета., 2007. - 480 с.:ил.
5. Курчанов, Н.А. Генетика человека с основами общей генетики. / Н.А. Курчанов. - СПб.: СпецЛит, 2006. - 174 с.
6. Мамонтов С.Г. Биология - М., 2004
7. Шевченко В.А., Топорнина Н.А., Стволинская Н.С. Генетика человека: Учеб.для студ. Высш. учеб. заведений. - М.: ВЛАДОС, 2002. - 240 с.9.
8. Ярыгин В.Н, В.И. Васильева, И.Н. Волков, В.В. Синелыцикова Биология. В 2 кн.: Учеб.для медиц. спец. Вузов М.: Высш. шк., 2003.— 432с.: ил.

Никола́й Петро́вич Дуби́нин Областью научных интересов Н. П. Дубинина была общая и эволюционная генетика, а также применение генетики в сельском хозяйстве. эволюционная генетика Вместе с А. С. Серебровским показал дробимость гена, а также явление комплементарности гена.А. С. Серебровскимгена комплементарности Опубликовал ряд важных научных работ по структуре и функциям хромосом, показал наличие в популяциях генетического груза летальных и сублетальных мутаций.хромосом генетического груза мутаций Также работал в области космической генетики, над проблемами радиационной генетики.радиационной

Дрейф генов как фактор эволюции Благодаря дрейфу частоты аллелей могут случайно меняться в локальных популяциях, пока они не достигнут точки равновесия – утери одного аллеля и фиксации другого. В разных популяциях гены «дрейфуют» независимо. Таким образом, дрейф генов ведет с одной стороны к уменьшению генетического разнообразия внутри популяций, а с другой стороны - к увеличению различий между популяциями, к их дивергенции по ряду признаков. Эта дивергенция в свою очередь может служить основой для видообразования.

Дрейф генов как фактор эволюции При высокой интенсивности отбора и высокой численности популяций влияние случайных процессов на динамику частот генов в популяциях становится пренебрежимо малым. Наоборот, в малых популяциях при небольших различиях по приспособленности между генотипами дрейф генов приобретает решающее значение. В таких ситуациях менее адаптивный аллель может зафиксироваться в популяции, а более адаптивный может быть утрачен. Аллель, утраченный в результате дрейфа, может возникать вновь и вновь за счет мутирования. Поскольку дрейф генов – ненаправленный процесс, то одновременно с уменьшением разнообразия внутри популяций, он увеличивает различия между локальными популяциями. Этому противодействует миграция. Если в одной популяции зафиксирован аллель А, а в другой а, то миграция особей между этими популяциями приводит к тому, что внутри обеих популяций вновь возникает аллельное разнообразие.

Популяционные волны и дрейф генов Численность популяций редко остается постоянной во времени. За подъемами численности следуют спады. С.С.Четвериков одним из первых обратил внимание на периодические колебания численности природных популяций, популяционные волны играют очень важную роль в эволюции популяций.

Серге́й Серге́евич Четверико́в () выдающийся русский биолог, генетик- эволюционист, сделавший первые шаги в направлении синтеза менделевской генетики и эволюционной теории Ч. Дарвина. Он раньше других ученых организовал экспериментальное изучение наследственных свойств у естественных популяций животных. Эти исследования позволили ему стать основоположником современной эволюционной генетики генетик эволюционист

Популяционные волны и дрейф генов В периоды резкого спада численности роль дрейфа генов сильно возрастает. В такие моменты он может становиться решающим фактором эволюции. В период спада частота определенных аллелей может резко и непредсказуемо меняться. Может происходить утеря тех или иных аллелей и резкое обеднение генетического разнообразия популяций. Потом, когда численность популяции начинает возрастать, популяция будет из поколения в поколение воспроизводить ту генетическую структуру, которая установилась в момент прохождения через «бутылочное горлышко» численности.



Эффект бутылочного горлышка в реальных популяциях Пример: Ситуация с гепардами – представителями кошачьих. Ученые обнаружили, что генетическая структура всех современных популяций гепардов очень сходна. При этом генетическая изменчивость внутри каждой из популяций крайне низка. Эти особенности генетической структуры популяций гепардов можно объяснить, если предположить, что относительно недавно данный вид прошел через очень узкое горлышко численности, и все современные гепарды являются потомками нескольких (по подсчетам американских исследователей, 7) особей.

Современный пример действия эффекта бутылочного горлышка популяция сайгака. Численность антилопы сайгак сократилась на 95 % от приблизительно 1 миллиона в 1990 году до менее чем в 2004, главным образом по причинам браконьерства для нужд традиционной китайской медицинысайгака сайгак1990 году 2004

Год Популяция американского бизона до особей особей особей

Эффект основателя Животные и растения, как правило, проникают на новые для вида территории относительно малыми группами. Частоты аллелей таких группах могут значительно отличаться от частот этих аллелей в исходных популяциях. За вселением на новую территорию следует увеличение численности колонистов. Возникающие многочисленные популяции воспроизводит генетическую структуру их основателей. Это явление американский зоолог Эрнст Майр, один их основоположников синтетической теории эволюции, назвал эффектом основателя.

Ясно, что основатели представляли собой очень маленькие выборки из родительских популяций и частоты аллелей в этих выборках могли сильно отличаться. Именно эффект основателя объясняет удивительно разнообразие океанических фаун и флор и обилие эндемичных видов на островах. Эффект основателя сыграл важную роль и в эволюции человеческих популяций. Обратите внимание, что аллель В (по системе групп крови АВ0) полностью отсутствует у американских индейцев и у аборигенов Австралии. Эти континенты были заселены небольшими группами людей. В силу чисто случайных причин среди основателей этих популяций могло не оказаться ни одного носителя аллеля В. Естественно, этот аллелей отсутствует и в производных популяциях.



Дрейф генов и молекулярные часы эволюции Конечным результатом дрейфа генов является полное устранение одного аллеля из популяции и закрепление (фиксация) в ней другого аллеля. Чем чаще тот или иной аллель встречается в популяции, тем выше вероятность его фиксации вследствие дрейфа генов. Расчеты показывают, что вероятность фиксации нейтрального аллеля равна его частоте в популяции.

Закономерность Большие популяции недолго «ждут» мутационного возникновения нового аллеля, но долго его фиксируют. Малые популяции очень долго «ждут» возникновения мутации, но после того, как она возникла, она может быть быстро зафиксирована. Из этого следует парадоксальный на первый взгляд вывод: вероятность фиксации нейтральных аллелей зависит только от частоты их мутационного возникновения и не зависит от численности популяций.

Закономерность Чем больше времени прошло с момента выделения двух видов из общего предкового вида, тем больше нейтральных мутационных замен различают эти виды. На этом принципе строится метод «молекулярных часов эволюции» - определения времени, прошедшего с момента, когда предки разных систематических групп стали эволюционировать независимо друг от друга.

Закономерность Американские исследователи Э. Цукуркендл и Л.Поллинг впервые обнаружили, что количество различий в последовательности аминокислот в гемоглобине и цитохроме с у разных видов млекопитающих тем больше, чем раньше разошлись их эволюционные пути.


Дрейф генов как фактор эволюции.
Дрейф генов, или генетико-автоматические процессы, - это явление ненаправленного изменения частот аллельных вариантов генов в популяции.
Мы можем рассматривать дрейф генов как один из факторов эволюции популяций. Благодаря дрейфу частоты аллелей могут случайно меняться в локальных популяциях, пока они не достигнут точки равновесия – утери одного аллеля и фиксации другого. В разных популяциях гены «дрейфуют» независимо. Поэтому результаты дрейфа оказываются разными в разных популяциях – в одних фиксируется один набор аллелей, в других – другой. Таким образом, дрейф генов ведет с одной стороны к уменьшению генетического разнообразия внутри популяций, а с другой стороны - к увеличению различий между популяциями, к их дивергенции по ряду признаков. Эта дивергенция в свою очередь может служить основой для видообразования. В ходе эволюции популяций дрейф генов взаимодействует с другими факторами эволюции, прежде всего с естественным отбором. Соотношение вкладов этих двух факторов зависит как от интенсивности отбора, так и от численности популяций. При высокой интенсивности отбора и высокой численности популяций влияние случайных процессов на динамику частот генов в популяциях становится пренебрежимо малым. Наоборот, в малых популяциях при небольших различиях по приспособленности между генотипами дрейф генов приобретает решающее значение. В таких ситуациях менее адаптивный аллель может зафиксироваться в популяции, а более адаптивный может быть утрачен. Как мы уже знаем, наиболее частым последствием дрейфа генов является обеднение генетического разнообразия внутри популяций за счет фиксации одних аллелей и утраты других. Мутационный процесс, напротив, приводит к обогащению генетического разнообразия внутри популяций. Аллель, утраченный в результате дрейфа, может возникать вновь и вновь за счет мутирования. Поскольку дрейф генов – ненаправленный процесс, то одновременно с уменьшением разнообразия внутри популяций, он увеличивает различия между локальными популяциями. Этому противодействует миграция. Если в одной популяции зафиксирован аллель А, а в другой а Эффект бутылочного горлышка В по системе групп крови АВ0
Популяционные волны и дрейф генов.
Эффект основателя. эффектом основателя В В
Дрейф генов и молекулярные часы эволюции. Расчеты показывают, что вероятность фиксации нейтрального аллеля равна его частоте в популяции. Каждый аллель из тех, что мы наблюдаем в популяциях, когда-то возник в результате мутации. Мутации происходят со средней частотой 10-5 на ген на гамету на поколение. Следовательно, чем меньше популяция, тем меньше вероятность, что в каждом поколении хотя бы одна особь в этой популяции окажется носителем новой мутации. В популяции, состоящей из 100000 особей, в каждом новом поколении с вероятностью близкой к единице найдется новый мутантный аллель, но частота его в популяции (1 на 200000 аллелей) и, следовательно, вероятность его фиксации будет очень низкой. Вероятность того, что эта же мутация в том же поколении возникнет у хотя бы одной особи в популяции, состоящей из 10 особей, ничтожно мала, но если такая мутация все же произойдет в этой популяции, то частота мутантного аллеля (1 на 20 аллелей) и шансы на его фиксацию будут относительно высокими. Большие популяции недолго «ждут» мутационного возникновения нового аллеля, но долго его фиксируют, а малые популяции очень долго «ждут» возникновения мутации, но после того, как она возникла, она может быть быстро зафиксирована. Из этого следует парадоксальный на первый взгляд вывод: вероятность фиксации нейтральных аллелей зависит только от частоты их мутационного возникновения и не зависит от численности популяций. метод «молекулярных часов эволюции» - определения времени, прошедшего с момента, когда предки разных систематических групп стали эволюционировать независимо друг от друга. с
Дрейф генов, или генетико-автоматические процессы, - это явление ненаправленного изменения частот аллельных вариантов генов в популяции.
Мы можем рассматривать дрейф генов как один из факторов эволюции популяций. Благодаря дрейфу частоты аллелей могут случайно меняться в локальных популяциях, пока они не достигнут точки равновесия – утери одного аллеля и фиксации другого. В разных популяциях гены «дрейфуют» независимо. Поэтому результаты дрейфа оказываются разными в разных популяциях – в одних фиксируется один набор аллелей, в других – другой. Таким образом, дрейф генов ведет с одной стороны к уменьшению генетического разнообразия внутри популяций, а с другой стороны - к увеличению различий между популяциями, к их дивергенции по ряду признаков. Эта дивергенция в свою очередь может служить основой для видообразования. В ходе эволюции популяций дрейф генов взаимодействует с другими факторами эволюции, прежде всего с естественным отбором. Соотношение вкладов этих двух факторов зависит как от интенсивности отбора, так и от численности популяций. При высокой интенсивности отбора и высокой численности популяций влияние случайных процессов на динамику частот генов в популяциях становится пренебрежимо малым. Наоборот, в малых популяциях при небольших различиях по приспособленности между генотипами дрейф генов приобретает решающее значение. В таких ситуациях менее адаптивный аллель может зафиксироваться в популяции, а более адаптивный может быть утрачен. Как мы уже знаем, наиболее частым последствием дрейфа генов является обеднение генетического разнообразия внутри популяций за счет фиксации одних аллелей и утраты других. Мутационный процесс, напротив, приводит к обогащению генетического разнообразия внутри популяций. Аллель, утраченный в результате дрейфа, может возникать вновь и вновь за счет мутирования. Поскольку дрейф генов – ненаправленный процесс, то одновременно с уменьшением разнообразия внутри популяций, он увеличивает различия между локальными популяциями. Этому противодействует миграция. Если в одной популяции зафиксирован аллель А
, а в другой а
, то миграция особей между этими популяциями приводит к тому, что внутри обеих популяций вновь возникает аллельное разнообразие. Эффект бутылочного горлышка
сыграл, по-видимому, очень значительную роль в эволюции популяций человека. Предки современных людей в течение десятков тысяч лет расселялись по всему миру. На этом пути, множество популяций полностью вымирало. Даже те, которые уцелели, часто оказывались на грани вымирания. Их численность падала до критического уровня. Во время прохождения через «бутылочное горлышко» численности частоты аллелей менялись по-разному в разных популяциях. Определенные аллели утрачивались полностью в одних популяциях и фиксировались в других. После восстановления численности популяций их измененная генетическая структура воспроизводилась из поколения в поколение. Эти процессы, по-видимому, и обусловили, то мозаичное распределение некоторых аллелей, которое мы сегодня наблюдаем в локальных популяциях человека. Ниже представлено распределение аллеля В
по системе групп крови АВ0
у людей. Значительные отличия современных популяций друг от друга могут отражать последствия дрейфа генов, который происходил в доисторические времена в моменты прохождения предковых популяций через «бутылочное горлышко» численности.
Популяционные волны и дрейф генов.
Численность популяций редко остается постоянной во времени. За подъемами численности следуют спады. С.С.Четвериков одним из первых обратил внимание на периодические колебания численности природных популяций, популяционные волны. Они играют очень важную роль в эволюции популяций. Дрейф генов мало сказывается на частотах аллелей в многочисленных популяциях. Однако в периоды резкого спада численности его роль сильно возрастает. В такие моменты он может становиться решающим фактором эволюции. В период спада частота определенных аллелей может резко и непредсказуемо меняться. Может происходить утеря тех или иных аллелей и резкое обеднение генетического разнообразия популяций. Потом, когда численность популяции начинает возрастать, популяция будет из поколения в поколение воспроизводить ту генетическую структуру, которая установилась в момент прохождения через «бутылочное горлышко» численности. Примером могут служить ситуация с гепардами – представителями кошачьих. Ученые обнаружили, что генетическая структура всех современных популяций гепардов очень сходна. При этом генетическая изменчивость внутри каждой из популяций крайне низка. Эти особенности генетической структуры популяций гепардов можно объяснить, если предположить, что относительно недавно (пару сотен лет назад) данный вид прошел через очень узкое горлышко численности, и все современные гепарды являются потомками нескольких (по подсчетам американских исследователей, 7) особей.
Эффект основателя.
Животные и растения, как правило, проникают на новые для вида территории (на острова, на новые континенты) относительно малыми группами. Частоты тех или иных аллелей таких группах могут значительно отличаться от частот этих аллелей в исходных популяциях. За вселением на новую территорию следует увеличение численности колонистов. Возникающие многочисленные популяции воспроизводит генетическую структуру их основателей. Это явление американский зоолог Эрнст Майр, один их основоположников синтетической теории эволюции, назвал эффектом основателя
. Эффект основателя играл, по-видимому, ведущую роль в формировании генетической структуры видов животных и растений, населяющих вулканические и коралловые острова. Все эти виды происходят от очень небольших групп основателей, которым посчастливилось достигнуть островов. Ясно, что эти основатели представляли собой очень маленькие выборки из родительских популяций и частоты аллелей в этих выборках могли сильно отличаться. Вспомним наш гипотетический пример с лисицами, которые, дрейфуя на льдинах, попадали на необитаемые острова. В каждой из дочерних популяций частоты аллелей резко отличались друг от друга и от родительской популяции. Именно эффект основателя объясняет удивительно разнообразие океанических фаун и флор и обилие эндемичных видов на островах. Эффект основателя сыграл важную роль и в эволюции человеческих популяций. Обратите внимание, что аллель В
полностью отсутствует у американских индейцев и у аборигенов Австралии. Эти континенты были заселены небольшими группами людей. В силу чисто случайных причин среди основателей этих популяций могло не оказаться ни одного носителя аллеля В
. Естественно, этот аллелей отсутствует и в производных популяциях.
Дрейф генов и молекулярные часы эволюции.
Конечным результатом дрейфа генов является полное устранение одного аллеля из популяции и закрепление (фиксация) в ней другого аллеля. Чем чаще тот или иной аллель встречается в популяции, тем выше вероятность его фиксации вследствие дрейфа генов. Расчеты показывают, что вероятность фиксации нейтрального аллеля равна его частоте в популяции.
Каждый аллель из тех, что мы наблюдаем в популяциях, когда-то возник в результате мутации. Мутации происходят со средней частотой 10 -5 на ген на гамету на поколение. Следовательно, чем меньше популяция, тем меньше вероятность, что в каждом поколении хотя бы одна особь в этой популяции окажется носителем новой мутации. В популяции, состоящей из 100000 особей, в каждом новом поколении с вероятностью близкой к единице найдется новый мутантный аллель, но частота его в популяции (1 на 200000 аллелей) и, следовательно, вероятность его фиксации будет очень низкой. Вероятность того, что эта же мутация в том же поколении возникнет у хотя бы одной особи в популяции, состоящей из 10 особей, ничтожно мала, но если такая мутация все же произойдет в этой популяции, то частота мутантного аллеля (1 на 20 аллелей) и шансы на его фиксацию будут относительно высокими. Большие популяции недолго «ждут» мутационного возникновения нового аллеля, но долго его фиксируют, а малые популяции очень долго «ждут» возникновения мутации, но после того, как она возникла, она может быть быстро зафиксирована. Из этого следует парадоксальный на первый взгляд вывод: вероятность фиксации нейтральных аллелей зависит только от частоты их мутационного возникновения и не зависит от численности популяций.
Поскольку частоты возникновения нейтральных мутаций примерно одинаковы у разных видов, то и скорость фиксации этих мутаций должна быть примерно одинаковой. Отсюда следует, что число мутаций, накопленных в одном и том же гене, должно быть пропорционально времени независимой эволюции этих видов. Иными словами, чем больше времени прошло с момента выделения двух видов из общего передкового вида, тем больше нейтральных мутационных замен различают эти виды. На этом принципе строится метод «молекулярных часов эволюции» - определения времени, прошедшего с момента, когда предки разных систематических групп стали эволюционировать независимо друг от друга.
Американские исследователи Э. Цукуркендл и Л.Поллинг впервые обнаружили, что количество различий в последовательности аминокислот в гемоглобине и цитохроме с
у разных видов млекопитающих тем больше, чем раньше разошлись их эволюционные пути. В дальнейшем эта закономерность была подтверждена на огромном экспериментальном материале, включающем десятки разных генов и сотни видов животных, растений и микроорганизмов. Оказалось, что молекулярные часы идут, как и следует из теории дрейфа генов, с постоянной скоростью. Калибровка молекулярных часов производится для каждого гена в отдельности, поскольку разные гены могут различаться по частоте возникновения нейтральных мутаций. Для этого оценивают количество замен накопленных в определенном гене у представителей таксонов, время дивергенции которых надежно установлено по палеонтологическим данным. После того, как молекулярные часы откалиброваны, их можно использовать для того, чтобы измерять время дивергенции между разными таксонами, даже в том случае, когда их общий предок пока не обнаружен в палеонтологической летописи. 1. Почему популяционные волны численности усиливают эффекты дрейфа генов? 2. Какую роль играет дрейф генов в формировании островных фаун и флор? 3. Объясните принцип молекулярных часов эволюции и его применение в эволюционных исследованиях.






